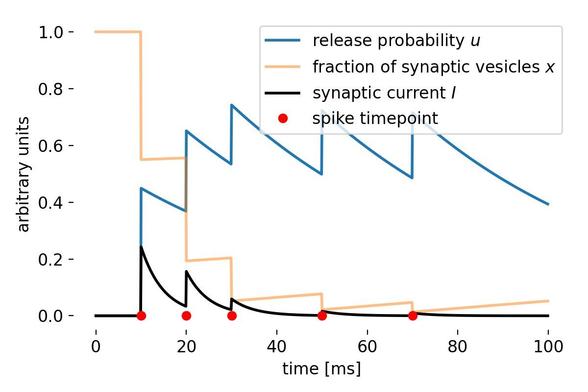
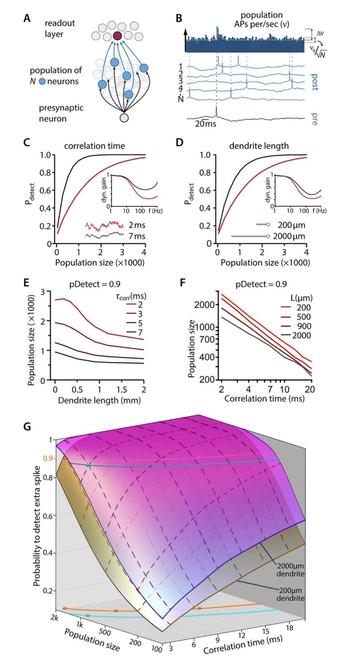
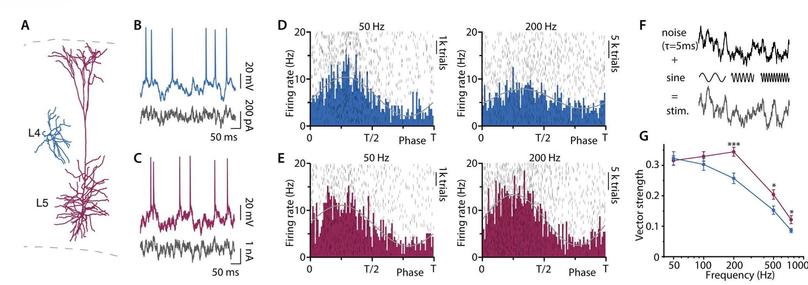
https://www.fogolf.com/1277256/which-spikes-shoes-should-fast-bowlers-wear/ Which Spikes Shoes Should Fast Bowlers Wear? #BOWLERS #FAST #GolfEquipment #GolfEquipmentVideos #GolfEquipmentVlog #GolfEquipmentYouTube #GolfShoes #GolfShoesVideos #GolfShoesVlog #GolfShoesYouTube #shoes #SpikedShoes #SpikedShoesVideos #SpikedShoesVlog #SpikedShoesYouTube #SPIKES #wear